1.子房不完全(3或)4室,具1下垂胚珠每室(如果2,同则1枚早期败育的),(6-)8肋,装饰,与其它相比较中部肋较少清楚凸起;
2.胚珠着生在二室或多室子房的中轴上。
3.花粉数量与胚珠比(P/O)为4000左右,杂交指数(OCI)介于3或4,繁育系统为异交型,需要传粉者;
4.胚珠来自高飞的树已网络以及漫衍遍地,并且它不长酿成扮饰物在家子公园。
5.的开花物侯、开花动态、访花者的种类、行为和访花频率,并对其繁育系统、花粉-胚珠比(P/O)、花粉活力进行了检测。
6.完整接种的早脆王胚珠随其成熟度的增加褐化加重,但切开远胚端接种金丝丰和冬枣的幼嫩胚珠更容易褐化。
7.对龙眼胚胎发育过程中胚珠内核酸、蛋白质和淀粉含量的变化进行了研究。
8.产生雌配子的雌配子体在大孢子囊即胚珠的珠心处发育。
9.雄蕊10,二体,花药同型,子房具柄,有胚珠多颗,花柱内弯;
10.芍药的结实率较低,野生居群平均只有23.27%的胚珠发育成种子。
11.取出早脆王胚珠,剥去珠被接种可以有效防止褐化。
12.用棉花胚珠切块诱导愈伤组织,经悬浮振荡培养、漂浮培养、滤纸桥法等方法诱导成纤维细胞。
13.假心皮化雌蕊上部变成近似倒U形的柱头结构区, 下部是完全突变的半开裂心皮, 其上着生4~14个不等的裸露的幼小胚珠;
14.结果表明,1)枇杷的胚珠为倒生胚珠,双珠被,核型胚乳;
15.金花茶子房3—5室,每室3—5个着生在中轴胎座上的具双珠被的倒生胚珠;
16.番茄的胚珠倒生。
17.结果表明,倒生的棉花胚珠可以分为合点区、株柄顶区、腰区和珠孔区。
18.胚珠为倒生型,具单珠被和薄珠心。
19.例如自交不亲和的腋花矮牵牛用一个子房的胎座上的全部胚珠进行离体培养和授以自花花粉。
20.绿豆子厉由单心皮构成,1室,有9—13个弯生胚珠着生在边缘胎座上。
21.细胞学研究发现,杂种胚阶段胚或胚珠的败育是冷杉种间杂交不成功的主要原因。
22.芙蕖的胚珠有涩味是收敛剂,有好处于人体的器官脾,肾,以及心脏。
23.花粉经常会被风吹到其他树上,使它们的胚珠受精。
24.虫媒传粉的植物平均每个胚珠生产约3450粒花粉,风媒传粉的植物生产的花粉几乎是这个数量的十倍。
25.运用常规石蜡制片法,在光学显微镜下观察了多糖在革苞菊胚珠发育过程中的消长动态。
26.统计3个居群柱头上落置的花粉量,76.58%以上的柱头接受了花粉,但柱头上的花粉量平均低于胚珠数;
27.其中一株开始生长出管状器官,用来传送精粉到授粉端花房的胚珠。
28.它由子房(内含一个或多个胚珠),花柱(部分植物没有花柱)和一个末端的接受表面-柱头构成。
29.子房具胚珠20—23枚,胚珠横生,珠被二层,薄珠心,孢原细胞直接起大孢子母细胞作用。
30.吉祥草花器官发生发育过程中出现雌蕊群发育停滞现象,以及吉祥草在小孢子及胚珠发育过程中出现败育情况,可能是吉祥草属结实率低的重要原因。
31.商陆属植物的胚珠维管束明显地来自花托的维管组织。
32.1986年美利坚合众国闻名杂志纽约客以五页刊登范光陵, 小余松, 拉地卡等主理之世界骚全国人民代表大会会及世界骚人与文化大会,并刊登范博士之新古诗 “胚珠” 为论断。
33.某些自交不亲和植物的花粉管能到达胚珠,但胚珠败育,这其中有的是由于受精失败,有的是由于受精后的早期胚珠败育。
34.子房、胚珠和胚培养、染色体加倍可用来克服远缘杂种的不育性。
35.被子植物不同于针叶植物和其他的裸子植物,被子植物的胚珠由子房所包被,受精后直接发育为果实。
36.每一子房室内约可产生160-240枚胚珠。
37.子房、胚珠和胚培养以及体细胞杂交可用于克服远缘杂交的不亲和性和杂种的不育性,有利于拓宽种质资源。
38.茉莉罕见的结实,是由极少数具胚囊胚珠的珠心组织填充空腔,而形成无繁殖功能的实心结构。
39.该文从雌配子体发育、胚珠发育及珠被发育等几个方面综述了近年来植物雌配子体发育的分子调控机制研究进展。
40.此后胚珠开始长大,五月中旬即分化形成珠被、珠心。
41.在植物,珠被在胚珠外形成一个封套。
42.以每胚珠鲜重计,DNA含量随胚珠的发育而逐渐增加。
43.本文首次报告了甜菜远缘杂交胚珠培养的结果。
44.授以油松花粉的红松,其胚珠发育至游离核时期即停止发育,与未授粉的红松胚珠发育很相似。
45.雄球花序中的不育胚珠和雄球花以及雌球花序中的胚珠均起源于环状总苞基部的分生组织。
46.进一步提高外源激素浓度,珠心细胞又可以分化出新的胚珠原基,进一步长大以后发育成许多新的胚珠。
47.扁桃果实由单心皮上位子房发育而成,边缘胎座,横生胚珠。
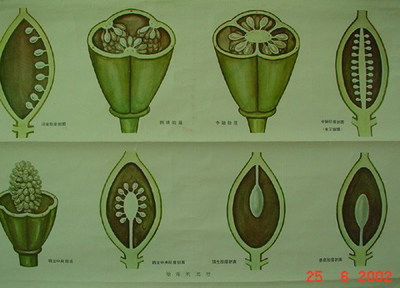
48.指在被子植物中着生胚珠的胎座位置。根据心皮数目的不同胎座式也有变化。
49.然而我们的遗传分析表明MADS21 对雄蕊、心皮和胚珠的身份决定没有作用。
50.最后,观测了五脉山黧豆的开花动态,并用杂交指数(OCI)、花粉-胚珠比(P/O)方法测定了其有性繁育系统类型。
51.加倍的MS培养基可以促进早脆王幼胚褐化,附加活性炭可以减轻早脆王完整胚珠褐化。
52.开花前后可以由于外施IAA,明显增加胚珠的纤维数量。
53.赤毒酸(GA_3)或GA_3加吲哚乙酸(IAA)能促进受精或未受精胚珠纤维的伸长。
54.在显微镜下观察发现,胎座末端的小细胞团分裂生长至受粉后43天具有胚珠的外型,之后胚珠开始延长,在受粉后99天逐渐形成成熟种子。
55.当胚发育最为充分而又尚未败育时取胚珠进行培养,存活胚的比例较高。
56.珠鳞基部近轴面着生2个胚珠;
57.不亲和的花粉管生长较慢,(至少在某些植物中)花粉管在到达胚珠之前就会破裂,多糖类胼胝质沉积在花粉管周围。
58.现在对后作用系统的了解不多,在后作用系统中,虽然花粉管到达胚珠,但胚珠败育。
59.被子植物通常的受精方式。花粉管通过珠孔进入胚珠。比较。
60.本文研究了“早玫瑰”和“新玫瑰”葡萄胚珠、胚乳和胚的发育。
61.本文研究了叶下珠科部分种类的花粉组织化学、花粉数和胚珠数,以及它们与传粉者之间的关系。
62.胚珠作为被子植物的有性生殖器官一直为植物学家所关注。
63.本文对拟南芥胚珠发育的分子调控机制进行了综述。
64.胚珠是研究器官形态发生和模式建成遗传分子机理的一个理想系统。
65.胚珠发育和雌配子体形成过程中遗传控制分子机理研究的材料主要来源于对突变体的发现和诱导。
66.研究还表明,该基因在胚珠和花瓣组织中也有较强的表达,但在其他组织中表达较弱或不表达。
67.被子植物子房中连接胚珠(之后的种子)和胎座的柄。
68.喙切可以大大提高金星无核胚珠的萌发率。
69.植物花粉中营养物质的储存形式以及单花的花粉数与胚珠数被认为与其传粉系统有一定的联系。
70.试管棉纤维主要是通过胚珠及愈伤组织诱导形成, 其影响因子主要是植物生长调节物质及外界环境。
71.亲和的花粉管到达胚珠,在胚囊中裂开后释放出精细胞。
72.花粉和胚珠——精子和卵子——长在同一棵树上但是具体部位不同。
73.基于返祖性畸形三胚珠球花的出现及其他形态特征,认为麻黄属胚珠数目减少是该属内物种特化的一个趋势;